Scientific research is maturing in a number of developing nations, which are trying to join North American and European nations as recognized centres of research. As recent stories show, the pressure to fulfill this vision--and to publish in English-language, international journals--has led to some large-scale schemes to commit academic fraud, in addition to cases of run-of-the-mill academic dishonesty.
In China, a widely-discussed incident involved criminals with a sideline in the production of fake journal articles, and even fake versions of existing medical journals in which authors could buy slots for their articles. China has been criticized for widespread academic problems for some time, for example, 2010 the New York Times published a report suggesting academic fraud (plagiarism, falsification or fabrication) was rampant in China and would hold the country back in its goal to become an important scientific contributor. In the other recent incident, four Brazilian medical journals were caught “citation stacking”, where each journal cited the other three excessively, thus avoiding notice for obvious journal self-citation, while still increasing their journal’s impact factor. These four journals were among 14 having their impact factors suspended for a year, with other possible incidences that were flagged but could not be proven involved Italian, a Chinese, and a Swiss journal.
There are some important facts that might provide context to these outbreaks of cheating. Both Brazil and China are nations where to be a successful scientist in the national system, you need to prove that you are capable of success on the world stage. This is probably a tall order in countries where scientific research has not traditionally had an international profile and most researchers do not speak English as their first language. In particular it leads to focus on values which are comparable across the globe, such as journal impact factors, as measures of success. In China, there is great pressure to publish in journals included on the Science Citation Index (SCI), a list of leading international journals. When researcher, department, and university success is quantified with impact factors and SCI publications, it becomes a numbers game, a GDP of research. Further, bonuses for publications in high caliber journals can double a poorly-paid researcher’s salary: a 2004 survey found that for nearly half of Chinese researchers, performance based pay was 50+ percent of their income. In Brazil, the government similarly emphasizes publications in Western journals as evidence of researcher quality.
It’s easy to dismiss these problems as specific to China or Brazil, and there are some aspects of the issue that are naturally country-specific. On the other hand, if you peruse Ivan Oransky’s Retraction Watch website, you’ll notice that academic dishonesty leading to article retraction is hardly restricted to researchers from developing countries. At the moment, the leading four countries in retractions due to fraud are the US, Germany, Japan, and then China, suggesting that Western science isn’t free from guilt. But in developing nations the conditions are ripe to produce fraud. Nationalistic ambition is funnelled into pressure on national scientists to succeed on the international stage; disproportionate focus on metrics of international success; high monetary rewards to otherwise poorly paid individuals for achieving these measures of success; combined with the reality that it is particularly difficult for researchers who were educated in a less competitive scientific system and who may lack English language skills, to publish in top journals. The benefits of success for these researchers are large, but the obstacles preventing their success are also huge. Combine that with a measure of success (impact factor, h-index) that is open to being gamed, and essentially relies on honesty and shared scientific principles, and it is not surprising that system fails.
Medical research was at the heart of both of these scandals, probably because the stakes (money, prestige) are high. Fortunately (or not) for ecology and evolutionary biology, the financial incentives for fraud are rather smaller, and thus organized academic fraud is probably less common. But the ingredients that seem to lead to these issues – national pressures to succeed on the world stage and difficulty in obtaining such success; combined with reliance on susceptible metrics – would threaten any field of science. And issues of language and culture are so rarely considered by English-language science (eg.), that it can be difficult for scientists from smaller countries to integrate into global academia. There are really two ways for the scientific community to respond to these issues of fraud and dishonesty – either treat these nations as second-class scientific citizens and assume their research may be unreliable, or else be available and willing to play an active role in their development. There are a number of ways the latter could happen. For example, some reputable national journals invite submissions from established international researchers to improve the visibility of their journals. In some nations (Portugal, Iceland, Czech Republic, etc), international scientists review funding proposals, so that an unbiased and external voice on the quality of work is provided. Indeed, the most hopeful fact is that top students from many developing nations attend graduate school in Europe and North America, and then return home with the knowledge and connections they gained. Obviously this is not a total solution, but we need to recognize fraud as problem affecting and interacting with all of academia, rather than solely an issue of a few problem nations.

Monday, September 30, 2013
Wednesday, September 25, 2013
Can common tradeoffs predict your supervisor’s functional type?
With Lanna Jin
1)

The invisible man/woman: The ultimate laissez-faire approach. You have the freedom to choose the ideas and projects that interest you, and the responsibility to make them work. Freedom can be replaced by frustrations when you need a signature, some support, or a manuscript commented on.
El generalissimo: "Here's my idea, now go do it." These labs are usually run in a top-down manner. Day to day operations are fairly hands off though, giving you room to work through those problems on your own. El generalissimo will reward their supporters well for good work.
The coach: The coach provides you the best of both worlds: enough rope to explore your ideas, but not so much to hang yourself. They are available for troubleshooting and brainstorming, but you ultimately have responsibility for your project. But if you rely on them to much, it's going to be hard to function without them.
The micromanager: These supervisors expect regular presence, frequent meetings, records of progress, and milestones to be met promptly. If you like working from home, leaving early or starting late, or need lots of freedom to be most productive it could be a poor fit. For students who thrive on structure and prefer set goals though, this might be an ideal environment.
2)

The superwoman/superman: This career superstar has made their name, possibly quite early, and they are passionate about their science. This can make for an exciting and successful lab experience, as new ideas and opportunities are always on the horizon. But since they have so many demands on their time, sometimes their capes (and they) are feeling a bit ragged.
The silverback: Labs of influential individuals can be an amazing opportunity for a graduate student. Silverback experiences might be quite variable, depending on how involved they are in day to day lab activities, their travel schedules, and the size of the lab. When they are available, they have a lot to teach a student about making a successful academic career.
Bad idea: This corner of the tradeoff (low interest in science, poor establishment in the career) probably doesn't exist in tenure-track faculty. If you do manage to find such a person, run away.
The unknown: A motivated but still unestablished supervisor is a blank slate. Their early career state means that they might have time and energy to devote to you and be especially motivated to see you and the lab succeed. On the other hand, they may not yet know how to manage people and their supervisor style could morph into anything - the coach, the micromanager, el generalissimo. A bit of a gamble.
Monday, September 23, 2013
Can intraspecific differences lead to ecosystem differences?
Sara Lindsay Jackrel and J. Timothy Wootton. 2013. Local adaptation of stream communities to intraspecific variation in a terrestrial ecosystem subsidy. Ecology. Online early.
It’s funny how complex outcomes can arise from simple realizations. For example, it is plausible that when there are differences among individuals of a species (like when local populations are adapted to the local environment), these could implications for function on the ecosystem scale. But while there is increasing evidence for the importance of intraspecific variation for ecological interactions within communities, the question of how intraspecific diversity scales up to ecosystem functioning is still ambiguous.
Sara Jackrel and Timothy Wootton explore this question in “Local adaptation of stream communities to intraspecific variation in a terrestrial ecosystem subsidy”. The basis for their study was simple: local adaptation is common, and populations/genotypes/ecotypes tend to be best adapted to the particular conditions of their locale. For example, “spatial variation in prey and predators can lead to a geographic mosaic of co-evolutionary interactions”. Further, these localized interactions can affect the greater ecosystem, if individuals or materials move between ecosystem boundaries.
In particular, the authors note that there is evidence that tree species composition riverside can alter the composition of the local aquatic community. This occurs via leaf litter fluxes into the river: the type and amount of leaf litter that falls into streams varies, and so the type of macroinvertebrates in the recipient stream also varies in response. These macroinvertebrates break down the leaf litter via shredding, collecting, and filtering, playing an important role in nutrient cycles. Leaf litter is carried from a given tree by wind or water and may decompose near or far away, creating a connection between ecosystems. The question then is whether macroinvertebrate compositional shifts will occur in response to intraspecific differences in leaf (i.e. trees), and what the implications might be for ecosystem functions such as leaf decomposition. To explore this question, Jackrel and Wootton performed reciprocal transplants of leaf litter material between eight sites along rivers in the Olympic Peninsula of Washington.
All eight of these sites were early successional forests dominated by red alder. The authors collected fresh leaves from alder trees, bagging leaves from each tree separately. These bags of leaves were either placed in the river adjacent to the trees they were taken from, or in a more distant site. Non-adjacent sites were either in the same river as the home site, or in different river all together. Leaf packs were weighed before and after spending 17-18 days in the river. This would allow comparison of how decomposition rates vary between home and away sites, and between home and away rivers.
Their results suggested a few interesting things. First, the identity of a tree affects the rate of decomposition of its leaves: individual alder trees’ leaves were highly variable in the rate of decomposition. Second, the combined identities of trees at a site seem to have affected the composition of the decomposer community at the home river site: put leaves from that site in another river with a new community of decomposers, and the decomposition rate drops significantly. In general, leaves decomposed significantly more rapidly when in their home river, regardless of whether at the home site or elsewhere along the river. But if they put leaves upstream from the home site, but in the same river, the rate of decomposition also dropped. Upstream decomposer communities were apparently much worse at breaking down leaves from novel communities of alders. However, if you put the leaves in sites downstream from home, the decomposition rates are not significantly different than in the home site. This is likely because of the directional movement of a river, such that downstream locations receive leaf litter from all upstream sites, and so downstream decomposer communities experience a greater variety of leaf litter than upstream sites. This might lead to upstream sites being more closely adapted to the individual trees in their neighbourhood than downstream sites, which receive inputs from a wide variety of trees. These results suggest that individual differences in trees at different spatial locations can matter, both locally, across trophic levels, and even across ecosystems.
Admittedly there is not a lot you can infer about the mechanisms at play from this preliminary experiment. One interesting follow up would be to measure compositional differences in aquatic macroinvertebrates at very fine scales in correspondence with differences in trees. Another important question is whether these communities differ via phenotypic plasticity, adaptation to local sites, or species sorting. But this paper does hint at one way in differences among individuals can shape local ecosystems and even structure distant ecosystems (e.g. downstream decomposer communities) through fluxes across boundaries. And that is a rather complicated implication from a logical and simple starting point.

It’s funny how complex outcomes can arise from simple realizations. For example, it is plausible that when there are differences among individuals of a species (like when local populations are adapted to the local environment), these could implications for function on the ecosystem scale. But while there is increasing evidence for the importance of intraspecific variation for ecological interactions within communities, the question of how intraspecific diversity scales up to ecosystem functioning is still ambiguous.
Sara Jackrel and Timothy Wootton explore this question in “Local adaptation of stream communities to intraspecific variation in a terrestrial ecosystem subsidy”. The basis for their study was simple: local adaptation is common, and populations/genotypes/ecotypes tend to be best adapted to the particular conditions of their locale. For example, “spatial variation in prey and predators can lead to a geographic mosaic of co-evolutionary interactions”. Further, these localized interactions can affect the greater ecosystem, if individuals or materials move between ecosystem boundaries.
In particular, the authors note that there is evidence that tree species composition riverside can alter the composition of the local aquatic community. This occurs via leaf litter fluxes into the river: the type and amount of leaf litter that falls into streams varies, and so the type of macroinvertebrates in the recipient stream also varies in response. These macroinvertebrates break down the leaf litter via shredding, collecting, and filtering, playing an important role in nutrient cycles. Leaf litter is carried from a given tree by wind or water and may decompose near or far away, creating a connection between ecosystems. The question then is whether macroinvertebrate compositional shifts will occur in response to intraspecific differences in leaf (i.e. trees), and what the implications might be for ecosystem functions such as leaf decomposition. To explore this question, Jackrel and Wootton performed reciprocal transplants of leaf litter material between eight sites along rivers in the Olympic Peninsula of Washington.
All eight of these sites were early successional forests dominated by red alder. The authors collected fresh leaves from alder trees, bagging leaves from each tree separately. These bags of leaves were either placed in the river adjacent to the trees they were taken from, or in a more distant site. Non-adjacent sites were either in the same river as the home site, or in different river all together. Leaf packs were weighed before and after spending 17-18 days in the river. This would allow comparison of how decomposition rates vary between home and away sites, and between home and away rivers.
Their results suggested a few interesting things. First, the identity of a tree affects the rate of decomposition of its leaves: individual alder trees’ leaves were highly variable in the rate of decomposition. Second, the combined identities of trees at a site seem to have affected the composition of the decomposer community at the home river site: put leaves from that site in another river with a new community of decomposers, and the decomposition rate drops significantly. In general, leaves decomposed significantly more rapidly when in their home river, regardless of whether at the home site or elsewhere along the river. But if they put leaves upstream from the home site, but in the same river, the rate of decomposition also dropped. Upstream decomposer communities were apparently much worse at breaking down leaves from novel communities of alders. However, if you put the leaves in sites downstream from home, the decomposition rates are not significantly different than in the home site. This is likely because of the directional movement of a river, such that downstream locations receive leaf litter from all upstream sites, and so downstream decomposer communities experience a greater variety of leaf litter than upstream sites. This might lead to upstream sites being more closely adapted to the individual trees in their neighbourhood than downstream sites, which receive inputs from a wide variety of trees. These results suggest that individual differences in trees at different spatial locations can matter, both locally, across trophic levels, and even across ecosystems.
Admittedly there is not a lot you can infer about the mechanisms at play from this preliminary experiment. One interesting follow up would be to measure compositional differences in aquatic macroinvertebrates at very fine scales in correspondence with differences in trees. Another important question is whether these communities differ via phenotypic plasticity, adaptation to local sites, or species sorting. But this paper does hint at one way in differences among individuals can shape local ecosystems and even structure distant ecosystems (e.g. downstream decomposer communities) through fluxes across boundaries. And that is a rather complicated implication from a logical and simple starting point.

Monday, September 16, 2013
Research in dangerous times
Research is full of disasters. Some are small – there’s nothing on the gel; someone published it first. Others are larger – ATVers just crushed the plants you were monitoring; you lost your lab notebook. But there are also real disasters – floods, fires, hurricanes, tornadoes, earthquakes.
The university where I am a postdoc, University of Colorado at Boulder, suffered through a 100-year flood this week. Though the university dealt with less water than many other parts of Boulder County, one-quarter of campus buildings have flood damage of some sort, there were power outages and the campus was closed and fairly inaccessible for several days. It sounds like my building got through okay – little to no water got in other than some roof leaks, and the power didn’t even go out. The water crept up into the parking lot, but never made it the rest of the way. But given the extent of the damage, undoubtably some other CU labs weren’t so fortunate. (And many people in Boulder County were not lucky at all).
What this experience reminded me of were the stories I had heard about university laboratories in New Orleans after Hurricane Katrina. The floodwaters there destroyed laboratories, took out research computers and servers, and killed 8000+ research animals in populations at Louisiana State University and Tulane University. This probably represents the loss of years of work and millions of dollars of funding. Data kept on damaged servers were gone. Research is generally so reliant on things we take for granted – like a reliable power source to maintain environmental conditions, and safe locations for irreplaceable data. When campuses are closed down, it may be impossible to reach laboratory populations of species that rely on regular care; routine measurements that may have been occurring for months or more are often disrupted; all the samples stored in a -80°C freezer might thaw out. And these are not uncommon outcomes when disasters hit universities. After Hurricane Sandy, criticisms were levelled at NYU for housing research animal populations in a basement, despite the potential for flooding. As a result, flooding killed thousands of mice, including strains representing a decade of research on forebrain development.
The personal impacts of these kind of losses are the saddest. The loss of months or years of data or experiments can be the nail in the coffin for grad students, who are reliant on time-limited funding as it is. A large enough setback may be the difference between finishing and dropping out for many students. Researchers, especially without tenure, face setbacks that could range between demoralizing and debilitating.
Like it or not, most predictions suggest disasters are going to be more common in the future. Often research labs rely on the disaster preparedness plans of the institution, and don't have specific plans for an individual lab. Every group (individual labs, departments, colleges, the institution as a whole) to some extent should consider disaster management plans. For most ecology labs, the considerations would not even be too onerous – consider how to maintain power to equipment that must remain on (e.g. freezers, experimental gear); ensure data is backed up in more than one physical location; if water damage is a possibility, be careful what you keep in the basement lab. Also, be sure to have contact information (including phone numbers) for all lab members - it's surprising how many labs neglect this - this way you can coordinate and ensure labmates and students are safe. Sometimes, no matter how prepared you are, loss and damage will be unavoidable. But hopefully, you are lucky and all of these preparations will be for nothing.

The university where I am a postdoc, University of Colorado at Boulder, suffered through a 100-year flood this week. Though the university dealt with less water than many other parts of Boulder County, one-quarter of campus buildings have flood damage of some sort, there were power outages and the campus was closed and fairly inaccessible for several days. It sounds like my building got through okay – little to no water got in other than some roof leaks, and the power didn’t even go out. The water crept up into the parking lot, but never made it the rest of the way. But given the extent of the damage, undoubtably some other CU labs weren’t so fortunate. (And many people in Boulder County were not lucky at all).
What this experience reminded me of were the stories I had heard about university laboratories in New Orleans after Hurricane Katrina. The floodwaters there destroyed laboratories, took out research computers and servers, and killed 8000+ research animals in populations at Louisiana State University and Tulane University. This probably represents the loss of years of work and millions of dollars of funding. Data kept on damaged servers were gone. Research is generally so reliant on things we take for granted – like a reliable power source to maintain environmental conditions, and safe locations for irreplaceable data. When campuses are closed down, it may be impossible to reach laboratory populations of species that rely on regular care; routine measurements that may have been occurring for months or more are often disrupted; all the samples stored in a -80°C freezer might thaw out. And these are not uncommon outcomes when disasters hit universities. After Hurricane Sandy, criticisms were levelled at NYU for housing research animal populations in a basement, despite the potential for flooding. As a result, flooding killed thousands of mice, including strains representing a decade of research on forebrain development.
The personal impacts of these kind of losses are the saddest. The loss of months or years of data or experiments can be the nail in the coffin for grad students, who are reliant on time-limited funding as it is. A large enough setback may be the difference between finishing and dropping out for many students. Researchers, especially without tenure, face setbacks that could range between demoralizing and debilitating.
Like it or not, most predictions suggest disasters are going to be more common in the future. Often research labs rely on the disaster preparedness plans of the institution, and don't have specific plans for an individual lab. Every group (individual labs, departments, colleges, the institution as a whole) to some extent should consider disaster management plans. For most ecology labs, the considerations would not even be too onerous – consider how to maintain power to equipment that must remain on (e.g. freezers, experimental gear); ensure data is backed up in more than one physical location; if water damage is a possibility, be careful what you keep in the basement lab. Also, be sure to have contact information (including phone numbers) for all lab members - it's surprising how many labs neglect this - this way you can coordinate and ensure labmates and students are safe. Sometimes, no matter how prepared you are, loss and damage will be unavoidable. But hopefully, you are lucky and all of these preparations will be for nothing.

Wednesday, September 11, 2013
When enemies catch up – declining invasive impacts in hogweed?
Dostál P., Müllerová J., Pyšek P., Pergl J. & Klinerová T. (2013). The impact of an invasive plant changes over time. Ecology Letters, Early View.
Invasive species are a major ecological issue in this age of global connectivity. Many ecosystems are unrecognizable today after invasion, and invasive species have considerable impacts on agriculture, tourism, aquaculture, forestry, and native biodiversity. Some invasive species have been present in novel environments for decades or centuries, but many are more recent introductions. What we are still trying to understand is whether invasive species impacts are constant through time. Do ecosystems and communities adapt to invasive species, and how do they do so? Some examples of how the impacts of invasions can decline through time exist: for example, the efficacy of allelopathic chemicals produced by garlic mustard (e.g. Alliaria petiolata) may lessen as native species adapt. Another hypothesis is that specialized pathogens or herbivores, absent at the start of the invasion, will increase with time, reducing populations of invasive species and allowing the recovery of native species.
 |
| Do specialized pathogens find escaped species eventually? |

The researchers also collected soil samples from these sites. These samples were used in a common garden experiment to test whether soil pathogens that affected hogweed success might differ between the sites with different ages of invasion. Soil from the sites with different ages of invasion history was either sterilized to kill living pathogens, or left unsterilized. To these different types of soil (sterilized or not, 0, 11, 28, 42, or 48 years of invasion), they planted 1) 1 hogweed seedling, 2) a mix of native species seed, or 3) 1 hogweed seedling and the mix of native species together. They then looked at how well the hogweed seedlings survived.
 |
| Results from the observational data showing changes in native productivity and richness, and hogweed cover as a function of time since initial invasion. (Dostal et al. 2013) |
 |
| Hogweed showed decreases in biomass as the length of invasion history in the soil increased, but only if the soil was not sterilized. (Dostal et al. 2013) |
Dostal et al. (2013) suggest that for these sites in the Czech Republic, hogweed dieback due to increased specialist pathogen load will prove key for native species recovery. They make it clear that this is not the same as advocating for a lack of response to the invasion. Ignoring established sites is providing a source for seed for new sites, puts rare native species at risk, and may leave management concerns such as declining ecosystem functions untouched. The other issue is that density dependent, stabilizing processes like increases in specialist pathogens, may lead to boom and bust cycles – initial rapid invasion might be followed by declining invasive populations as specialist enemies increase, but declining invasive populations would lead to declines in specialist enemies, and increased invasive pressures could start anew. Such a situation wouldn’t lead to a general decline in invasive species, but rather a new (non-equilibrium) state for the community.
Thursday, September 5, 2013
The evolution of evolution, LEGO in the lab and other Science-y links
My week is coming to an early end as I head off to some friends' wedding tomorrow, so in lieu of another post, here are some interesting science links from around the internet this week :)
This infographic explores how thinking about evolution has changed since Darwin. It shows pretty clearly the circuitous path that science takes, the way ideas converge and diverge, and ultimately become more nuanced and complicated.

A theoretical physicist blogger answers the question "should you write a science blog?". She mentions the basic, but undeniably key points - do you have time? do you really have time? do you like writing? I also like her advice: "don't be afraid of your readers".
Mental illness can be exacerbated or first show up during grad school. Even in liberal academia, talking about mental health issues can be a bit taboo, something that doesn't help anyone. A blog post from Nash Turley considers the issues and implications.
Another serious issue, related to issues of gender in science: an article in the Economist presents evidence that women authors tend to be less cited than male authors, and this was in part due to less self-citation by women.
Also, LEGO now has its first female scientist character, and with her short hair, goggles, lab coat and gloves, she's a great lab safety role model too ;)

Of course, LEGO has many other science-related applications :)
Edit - the link about blogging was initially incorrect, should be correct now.
This infographic explores how thinking about evolution has changed since Darwin. It shows pretty clearly the circuitous path that science takes, the way ideas converge and diverge, and ultimately become more nuanced and complicated.

A theoretical physicist blogger answers the question "should you write a science blog?". She mentions the basic, but undeniably key points - do you have time? do you really have time? do you like writing? I also like her advice: "don't be afraid of your readers".
Mental illness can be exacerbated or first show up during grad school. Even in liberal academia, talking about mental health issues can be a bit taboo, something that doesn't help anyone. A blog post from Nash Turley considers the issues and implications.
Another serious issue, related to issues of gender in science: an article in the Economist presents evidence that women authors tend to be less cited than male authors, and this was in part due to less self-citation by women.
Also, LEGO now has its first female scientist character, and with her short hair, goggles, lab coat and gloves, she's a great lab safety role model too ;)

Of course, LEGO has many other science-related applications :)
Edit - the link about blogging was initially incorrect, should be correct now.
Tuesday, September 3, 2013
Studying Frankenstein: what can we learn from novel ecosystems?
There's been some discussion going around ecolog about an article telling the ecological story of Ascension Island. I should note that the original article is not a great example of science writing; it tries to create conflict that doesn’t exist and lacks a reasonable understanding of ecological theory. There are a couple linked chapters/publications about Ascension Island that make better additions to the story though (1, 2).
Ascension Island is one of those tiny islands first visited by Europeans in the 1600s. Like many young, small, isolated islands (1200 mi to the next nearest island), it was highly depauperate (~25-30 species of plants). Like many such islands, once humans became regular visitors, new species began to make their to way Ascension. The Brits and their love of cultivating and homogenizing particularly altered the island, and they systematically introduced species calculated to provide ecosystem services, aesthetic value, and food.
As a result, Ascension Island changed strikingly – once an island with lowland deserts and a rocky, barren mountainside, the mountain is today known as Green Mountain. The originally depauperate mountain is now lush with three different vegetation zones, a large variety of plants including “banana, ginger, juniper, raspberry, coffee, ferns, fig trees, Cape Yews, and Norfolk Island pines”, and a complex cloud forest. The original article presents this as some inexplicable outcome, but frankly it seems in keeping with existing ecological ideas. Under island biogeography, if you decrease the distance from an island to the mainland (including via human-aided dispersal), diversity should increase. Given the massive number of species that were introduced, and the coddling they received to aid their establishment, heightened diversity is hardly a surprise. And though the original article suggests that shared evolutionary history is necessary for complex ecosystems, coevolution is hardly a requirement for a functioning ecosystem to develop. Species may be able to coexist despite lacking a shared history--niches may not be filled as tightly as in a long-established, coevolved community, but invasive species research in general should have taught us that novel species combinations can easily occur. Secondly, many of the introduced species on the island are from the same part of the world and likely do share evolutionary history.
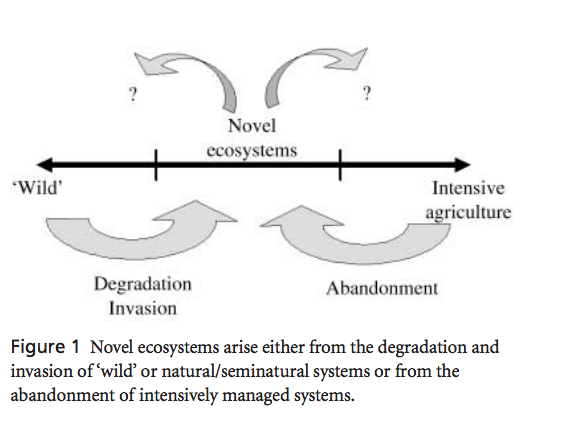
I hadn't given much thought before to the concept of “novel ecosystems” and it has received little attention from the ecological literature (excepting the odd papers, and much more attention from a conservation and management angle). Ascension is a particularly striking example of how human modification leads to ecosystems which are entirely different from anything that has ever been present on the planet. Novel ecosystems have been defined in a number of ways. Generally, they are synthetic ecosystems that include conditions and combinations of organisms never before in existence, and do not depend on human maintenance to persist (as agriculture fields would). Novel ecosystems may be considered to be the outcome of abandonment of human managed systems or else the degradation of existing systems through human activities and invasion (figure). Of course there are incredibly few ecosystems that aren’t affected in some way by human activities (especially in this age of intentional and unintentional human-mediated species introductions), but it is the truly unique ones that are particularly interesting.
There are at least two ways to approach novel ecosystems. One approach is parallel with invasive species and conservation research, and in fact these research areas overlap a fair amount. This is the way in which most research on novel ecosystems seems to be framed. Novel ecosystems carry many of the same issues about making value judgments as invasive species research, and issues of management and whether novel ecosystems can or should be returned to their original state dominate. For example, the conflict between maintaining alpha (island) and gamma (global) diversity exists on Ascension Island– modern, invaded Ascension Island provides greater diversity and ecosystem functioning (erosion control, food, temperature moderation, habitat) than the original barren landscape. But the original endemic species, not surprisingly, have gone extinct or are increasingly at risk.
But focusing solely on these difficult value-laden questions seems to have been at the cost of exploring the value of novel ecosystems as a study system. The most interesting examples of novel ecosystems are not simply modified or invaded ecosystems, but ecosystems that truly never existed before. Like post-shale dump landscapes in Scotland, where the refuse from mining is now host to unique grasslands that act as refugia for locally rare species; or the San Francisco Bay, which now is utterly unrecognizable compared to historical descriptions due to heavy invasion; or urban ecosystems with their unique habitats and issues; or even the habitat and connectivity created by stone fences which now occur on most continents. The questions here aren't always about invasion and management, but instead focus on what the new community looks like. How do novel communities assemble, what processes dominate (mass effects, environmental filtering, competition, predation, etc, etc)? How does ecosystem function relate to the community that assembles? Most BEF research after all, is focused on more traditional ecosystems. What leads to stability in a novel ecosystem, or are they stable at all? They can function is an example of highly unfortunate but also highly informative ‘natural’ experiments for ecologists. But at the moment, if you search for "novel ecosystems" on Google Scholar, the title words are "management", "conservation", "restoration" or "invasion". Actually, there probably are ecologists doing work on novel ecosystems from a purely ecological perspective, but this work gets grouped with disturbance, invasion, and urban ecology: it just remains to consider them in a more unified fashion. If the conversation remains focused only on the conservation issues (as the discussion on ecolog seemed to shift to rapidly), it just seems like we're limiting ourselves a little.
Ascension Island is one of those tiny islands first visited by Europeans in the 1600s. Like many young, small, isolated islands (1200 mi to the next nearest island), it was highly depauperate (~25-30 species of plants). Like many such islands, once humans became regular visitors, new species began to make their to way Ascension. The Brits and their love of cultivating and homogenizing particularly altered the island, and they systematically introduced species calculated to provide ecosystem services, aesthetic value, and food.
As a result, Ascension Island changed strikingly – once an island with lowland deserts and a rocky, barren mountainside, the mountain is today known as Green Mountain. The originally depauperate mountain is now lush with three different vegetation zones, a large variety of plants including “banana, ginger, juniper, raspberry, coffee, ferns, fig trees, Cape Yews, and Norfolk Island pines”, and a complex cloud forest. The original article presents this as some inexplicable outcome, but frankly it seems in keeping with existing ecological ideas. Under island biogeography, if you decrease the distance from an island to the mainland (including via human-aided dispersal), diversity should increase. Given the massive number of species that were introduced, and the coddling they received to aid their establishment, heightened diversity is hardly a surprise. And though the original article suggests that shared evolutionary history is necessary for complex ecosystems, coevolution is hardly a requirement for a functioning ecosystem to develop. Species may be able to coexist despite lacking a shared history--niches may not be filled as tightly as in a long-established, coevolved community, but invasive species research in general should have taught us that novel species combinations can easily occur. Secondly, many of the introduced species on the island are from the same part of the world and likely do share evolutionary history.
 |
| The mountain before and after. From Catling & Stroud. |
 |
| from Hobbs et al. 2006 |
There are at least two ways to approach novel ecosystems. One approach is parallel with invasive species and conservation research, and in fact these research areas overlap a fair amount. This is the way in which most research on novel ecosystems seems to be framed. Novel ecosystems carry many of the same issues about making value judgments as invasive species research, and issues of management and whether novel ecosystems can or should be returned to their original state dominate. For example, the conflict between maintaining alpha (island) and gamma (global) diversity exists on Ascension Island– modern, invaded Ascension Island provides greater diversity and ecosystem functioning (erosion control, food, temperature moderation, habitat) than the original barren landscape. But the original endemic species, not surprisingly, have gone extinct or are increasingly at risk.
But focusing solely on these difficult value-laden questions seems to have been at the cost of exploring the value of novel ecosystems as a study system. The most interesting examples of novel ecosystems are not simply modified or invaded ecosystems, but ecosystems that truly never existed before. Like post-shale dump landscapes in Scotland, where the refuse from mining is now host to unique grasslands that act as refugia for locally rare species; or the San Francisco Bay, which now is utterly unrecognizable compared to historical descriptions due to heavy invasion; or urban ecosystems with their unique habitats and issues; or even the habitat and connectivity created by stone fences which now occur on most continents. The questions here aren't always about invasion and management, but instead focus on what the new community looks like. How do novel communities assemble, what processes dominate (mass effects, environmental filtering, competition, predation, etc, etc)? How does ecosystem function relate to the community that assembles? Most BEF research after all, is focused on more traditional ecosystems. What leads to stability in a novel ecosystem, or are they stable at all? They can function is an example of highly unfortunate but also highly informative ‘natural’ experiments for ecologists. But at the moment, if you search for "novel ecosystems" on Google Scholar, the title words are "management", "conservation", "restoration" or "invasion". Actually, there probably are ecologists doing work on novel ecosystems from a purely ecological perspective, but this work gets grouped with disturbance, invasion, and urban ecology: it just remains to consider them in a more unified fashion. If the conversation remains focused only on the conservation issues (as the discussion on ecolog seemed to shift to rapidly), it just seems like we're limiting ourselves a little.
Wednesday, August 28, 2013
The species we’ve neglected
 |
| Species in last 3 months' papers in Ecology Letters. "Multiple species" tended to be meta-analyses. |
Browse the abstracts of a high profile ecological journal (for example, Ecology Letters, right) and one pattern you’ll notice is that high impact, hypothesis-driven ecology usually involves a small pool of focal species. Plants, for example, dominate any discussion of community ecology and have since Clements’ and Gleason’s arguments. It is not that hard to see why – plants don’t move, for one, live in speciose groups, and often complete a full lifecycle in a matter of months. They are also the lowest trophic level and so pesky multiple trophic level interactions can be omitted.
Establishing a new system may be a time-consuming activity with the possibility of failure. But these under-utilized species have something new to tell ecology. This is not to say that the popular systems of species have nothing to tell us anymore – not at all, given all the complexities of ecological dynamics – but they bias the story. The ecological processes at play are not likely much different between novel systems and traditional ones. But the same processes interact in different ways and differ in importance across systems, and so we may have unrealistic expectations about the importance of, say, competition, if we only focus on 1 or 2 systems. To follow Vellend’s (2011) framework, the processes of selection, drift, speciation, and dispersal are part of any ecological system. What differs is their importance, and their importance differs for reasons related to the ecological context and evolutionary history a species experiences. This is the reason that comparing Mark McPeek’s work on neutrality in damselflies with Jonathan Losos’ findings about adaptive radiation in anoles is so interesting. No one questions that adaptive radiations may drive one set of species and neutrality another, the real question is what about their contexts produces to this result. Unfortunately, if our current set of focal species is small, we are limited in our ability to make such informative comparisons.
Many of the limitations on species have been methodological: popular systems tend to involve amenable species. Other species may be very small, very mobile, very difficult to identify, or highly specialized in their habitats. This creates difficulties. But when we overcome them, the results are often revolutionary. For example, consider the current burst of interest in belowground interactions, once their incredible importance to plant community interactions became clear (e.g. Klironomos 2002, Nature). Further, techniques are continually improving in ways which make new systems tenable.
So we should continue to focus on a few well-understood systems, attempting to perfect our understanding and predictive abilities. There is much value in understanding a system as completely as possible. But on the other hand, we can limit ourselves by focusing too much. It seems like one of the big areas for growth in modern ecology is simply to expand into novel ecological systems.
(**It's probably too general and a bit unfair to refer to all plants and all insects as though they are monolithic groups, since they are each large and varied (which is part of the reason they've been useful thus far). And some of their great representation may in fact relate to the number of species available to study. But I do think the general point about the problem of focusing too much holds.**)
Monday, August 26, 2013
Everything you wanted to know about peer review (but no one mentions)
Since the British Ecological Society has published an introduction to reviewing successfully, here’s a short list of additional, less noted, observations about the reviewing process.
For example, excitement for reviewing is proportional to the number of reviews you have done
Manuscripts fall broadly into only a few categories. They might be deeply flawed and unpublishable, and therefore easy to review; or they might be uniformly excellent and therefore easy to review. But these are the least common types you will experience. Most manuscripts have both strengths and weaknesses and fall somewhere on the spectrum between “accept” and “reject”. These are the papers that take the most time, since you must weigh the flaws against the strengths, agonize over what changes to suggest, what suggestions might get them around the biggest issues, and what recommendation to give the editor. It’s also easy to fall into Monday morning quarterbacking and make impractical suggestions - why didn’t you design your experiment like this? Why didn’t you measure that? While these points might be reasonable and relevant, but it is important to be clear as to what is within the scope of a revision and what is a bigger picture problem.
Reviewing is of course an important service to ecology. It can also makes a number of subtle contributions to your own professional development. Once the novelty of someone caring about your opinion has worn off, the best part of reviewing may be things you don’t expect.
For example, excitement for reviewing is proportional to the number of reviews you have done
- When you are first asked, reviewing feels like a great honour. It is one of the first signs that some group larger than your lab or department recognizes your existence. You will spend an unreasonable amount of time perfecting your review.
 |
| This plot would not survive peer review. |
- The novelty will wear off, and your enthusiasm upon receiving a review request will decline, usually in relation to your increasing workload.
- Sadly, the urgent need to complete a review may also wane. You will probably submit the first review early, but after that…
Manuscripts fall broadly into only a few categories. They might be deeply flawed and unpublishable, and therefore easy to review; or they might be uniformly excellent and therefore easy to review. But these are the least common types you will experience. Most manuscripts have both strengths and weaknesses and fall somewhere on the spectrum between “accept” and “reject”. These are the papers that take the most time, since you must weigh the flaws against the strengths, agonize over what changes to suggest, what suggestions might get them around the biggest issues, and what recommendation to give the editor. It’s also easy to fall into Monday morning quarterbacking and make impractical suggestions - why didn’t you design your experiment like this? Why didn’t you measure that? While these points might be reasonable and relevant, but it is important to be clear as to what is within the scope of a revision and what is a bigger picture problem.
Reviewing is of course an important service to ecology. It can also makes a number of subtle contributions to your own professional development. Once the novelty of someone caring about your opinion has worn off, the best part of reviewing may be things you don’t expect.
- For example, one of the best parts of reviewing a paper in the same area as your research is seeing what literature the authors cite and how they cite them– some real gems you've missed can show up.
- Reviewing a paper that falls so exactly in your body of knowledge that you feel completely qualified is a great feeling. It’s nice to be reminded that you have (mostly) mastered a topic you care about.
- When you are asked to review a paper that combines some topic or method you are well-versed in with ideas or systems or methodologies you are not familiar with, it can be truly eye opening. The funnest papers to review are the ones where you think “I never thought of that!”.
- Reviewing can give you the clarity to recognize the weaknesses in your own work.
Thursday, August 22, 2013
Quotes that stick. #INT13
I'm back in Toronto now, and here are some quotes from talks that have really stuck with me. INTECOL was a great meeting, it was very interesting to hear about all the research from around the world. I hope all the attendees had a great time.
Sandra Diaz: “We just don't know enough to understand how
functional diversity links to environmental change and ecosystem services.”
Erika Edwards: “big phylogeny, big trait data set analyses
leave me feeling a little empty”
Erika Edwards: “carbon economy is part of the whole
organism, not single traits.”
Joel Cohen: “Mathematics is like sex, you can talk about it
but you shouldn't do it in public.”
Enrique Chaneton, Describing what happen during a study
looking at the effects of grazing on ecosystem decomposition rates: “A volcano
erupted during the study and sometimes shit happens, ….. the volcano killed
many of the cattle.”
Carsten Meyer, Talking about global data availability in
large databases: “Countries that under report are large emerging economies
(china, India, Brazil, Russia) which could finance these efforts but for some
reason do not.”
Ove Hoegh-Guldberg, ‘To get change we need to reach more
than the brain, but the human heart”
Subscribe to:
Posts (Atom)